
Table of Contents
-
Volume 56 Issue 6
Special issue: Biogeographic Patterns in the Pacific and Australasian Regions
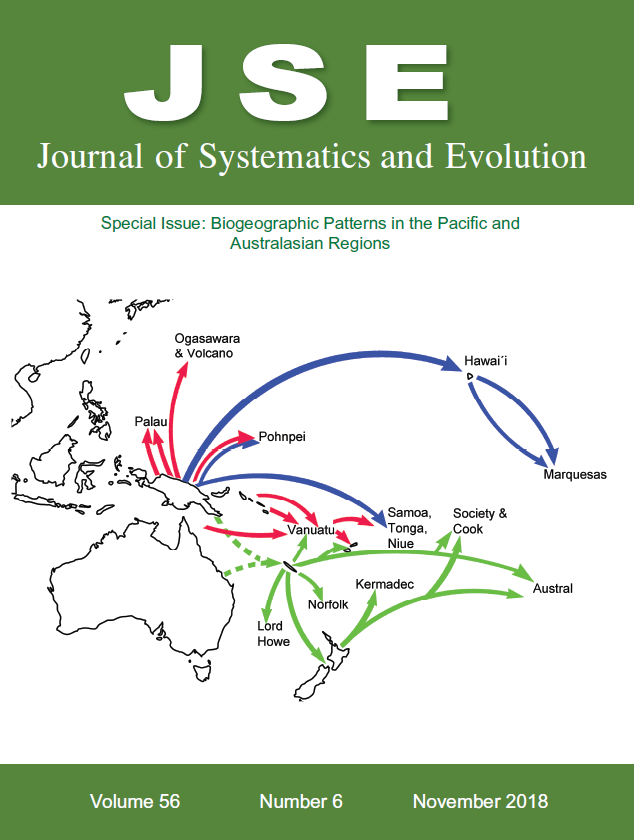
Cover illustration: Dispersal pattern of Melicope (Rutaceae, Citrus family) in the Pacifi c. Colors of arrows are according to the main phylogenetic lineages within the genus. See Appelhans et al., pp. 576–599 in this issue.
Editors-in-Chief
Song Ge
Jun Wen
Song Ge
Jun Wen
Impact Factor
3.7
JCR 2022 IF ranking: 60/238 (Plant Sciences, top 25%, Q2 quartile)
Journal Abbreviation: J Syst Evol
CN: 11-5779/Q
Frequency: Bi-monthly
Journal Abbreviation: J Syst Evol
| ISSN: | 1674-4918 (Print) 1759-6831 (Online) |
Frequency: Bi-monthly
Sponsors


Publisher
Co-Sponsor


Scan to view the journal on your mobile
device
device

Scan to follow us on WeChat