Cyperaceae are among the 10 most diverse angiosperm families and the third largest among monocots (Christenhusz & Byng, 2016), with a critical ecological role in wetlands and in high-latitude and high-elevation ecosystems. Sedges represent an ideal model plant family to study evolutionary biology because of their species richness, global distribution, large discrepancies in lineage diversity, broad range of ecological preferences, and adaptations including multiple origins of C4 photosynthesis and holocentric chromosomes (Larridon et al., 2021a).
In the last two decades, molecular phylogenetics has revolutionized our understanding of the sedge family. The first phylogenetic approaches beyond morphology used chromosomes (e.g., Heilbron, 1924), relationships with parasitic fungi (e.g., Savile & Calder, 1954), and flavonoids (e.g., Kukkonen, 1971). However, the current systematic framework of Cyperaceae started to take root with the advent of the first suprageneric phylogeny of Cyperaceae (based on a single chloroplast marker -rbcL- and a limited sampling; Muasya et al., 1998) and the first molecular phylogeny of Carex s.l. (based on nuclear -ITS- sequence data; Starr et al., 1999). Through collaboration and technical advances such as high-throughput sequencing, we have now achieved much more in-depth sampling of taxa and DNA regions. Thanks to these efforts, the latest evolutionary relationships retrieved for the sedge family have obtained stronger support than in previous works. On the one hand, new phylogenomic approaches have generated a wealth of sequence data, allowing re-investigation of fast-evolving Cyperaceae lineages in which relationships could not be resolved using Sanger sequencing data (Larridon et al., 2020). On the other hand, the increasing availability of bioclimatic data, digitized herbarium material, communication channels, plant material interchange opportunities, and new bioinformatic tools has allowed the gathering, analysis, and processing of information in a way that was not possible just a decade ago. This has enabled continuous and successful collaborations among researchers across the globe, and it has opened new research horizons for the international Cyperaceae research community to investigate the systematics, taxonomy, biogeography, ecology, morphology, and evolution of key characters in sedges such as chromosomes and C4 photosynthesis. The achievements of the Global Carex Group (GCG) serve as a best-practice example. In 2013, this consortium of researchers initiated a collaborative sampling effort that led to an on-hand sampling of about 80% of the approximately 2000 species of the giant genus Carex. Among other milestones, the GCG has conducted a massive sequencing effort (Jiménez-Mejías et al., 2016a), performed a thorough revision of the fossil record of Carex (Jiménez-Mejías et al., 2016b), and provided Carex with a robust backbone phylogeny of its major lineages using phylogenomics (Villaverde et al., 2020). Together, this allowed for the first time the possibility of addressing the biogeographic history of Carex from a global perspective using a sampling of >60% of the c. 2000 spp. of the genus (Martín-Bravo et al., 2019). Moreover, with the recent creation of the International Sedge Society, wider and even more ambitious research projects involving the entire international Cyperaceae research community will soon be possible. This Special Issue brings together a series of the most modern studies on the systematics and evolution of Cyperaceae that are the result of the new opportunities offered by our data-rich era and international collaboration.
The increased availability of DNA sequence data has led to higher-level taxonomic changes in Cyperaceae. Previously organized into four subfamilies, 14 tribes, and 104 genera (Goetghebeur, 1998), the earliest DNA sequence studies (e.g., Muasya et al., 2009a, 2009b) demonstrated that many of these traditional groups were defined by plesiomorphies and homoplasies, which meant that numerous tribes and genera were unnatural. At the tribal level, significant changes were made to the circumscription of tribe Schoeneae with the exclusion of lineages now placed in tribes Carpheae and Cladieae (Semmouri et al., 2019), and within the Scirpo-Caricoid Clade, where Scirpeae was narrowly defined and four new tribes were created (Léveillé-Bourret et al., 2018a, 2018b; Léveillé-Bourret & Starr, 2019). In this Special Issue, Starr et al. (2021) and Larridon et al. (2021a) use targeted sequencing data in concert with traditional data sources to propose new tribal rearrangements and re-circumscriptions. Starr et al. (2021) focus on tribe Fuireneae s.l., a group long known to be paraphyletic (e.g., Semmouri et al., 2019), but whose circumscription had not changed due to a lack of support for trees. With the benefit of significant novel sequence data and a topology congruent with morphology and embryo features, they propose a new classification for the Fuireneae s.l. Grade that involves four tribes, one of which is new. Larridon et al. (2021a) use a representative sampling of almost all the recognized sedge genera to provide a new classification of the family supported by phylogenomic data. This paper also provides a complete overview of the classification history of Cyperaceae.
Molecular phylogenetic studies have equally contributed to rearrangements at the generic level. Main changes to generic circumscriptions occurred in tribes Schoeneae (e.g., Elliott & Muasya, 2017; Larridon et al., 2018a, 2018b; Barrett et al., 2020, 2021a, 2021b), Cariceae (Global Carex Group, 2015), Abildgaardieae (e.g., Roalson et al., 2019; Larridon et al., 2021b), Trichophoreae (Léveillé-Bourret et al, 2020), and Cypereae (e.g., Larridon et al., 2011, 2014). In this Special Issue, using both a targeted sequencing and an nrDNA data set, Starr et al. (2021) also recircumscribe Schoenoplectus and Schoenoplectiella to be reciprocally monophyletic, two genera whose limits have never clearly been marked.
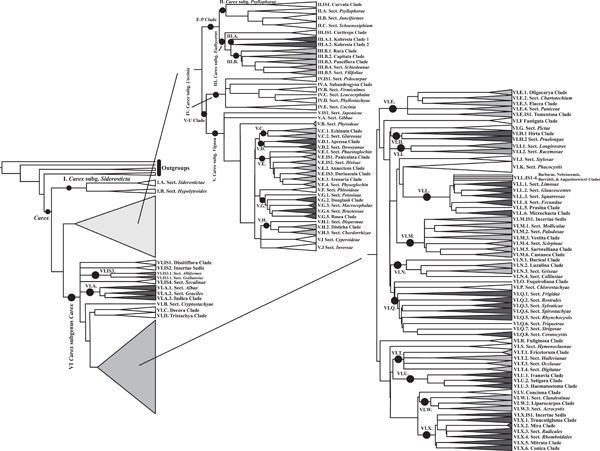
At the infrageneric level, the GCG re-examines the classification of the giant genus Carex. Being one of the three largest non-apomictic genera of the world, the revision and recircumscription of Carex started by merging it with its segregate genera to become monophyletic (GCG, 2015) and has continued through a series of later publications (e.g., Jiménez-Mejías et al., 2016; Villaverde et al., 2020). In this Special Issue, Roalson et al. (2021) re-evaluate the infrageneric classification of Carex using a systematic perspective and organizing principles that will serve as a roadmap to future research on Carex.
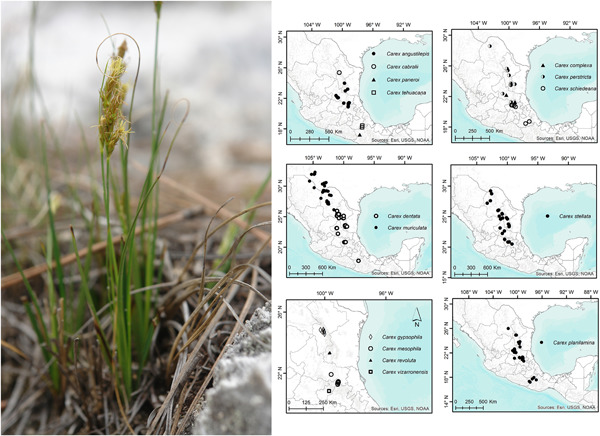
At lower taxonomic levels, much work remains to resolve relationships between species, uncover previously undescribed diversity, and unravel species complexes. Here, using an integrated taxonomy approach, Reznicek et al. (2021) monograph Carex sect. Schiedeanae, a highly diverse group in Mexico, where they describe 10 new species. In the same line, Lu et al. (2021) revise the phylogenetic relationships and morphological limits of two Carex sections whose delimitation has been unclear—sects. Confertiflorae and Molliculae—and provide the first insights into Carex phylogeography in Eastern Asia. These examples show the strength of combining multiple data sources to investigate the evolution and diversity of Cyperaceae lineages and it demonstrates that considerable work is still needed on understudied lineages and geographic regions, as also found by recent studies in other sedge groups (e.g., Jiménez-Mejías et al., 2021; Luceño et al., 2021). Using population genomics based on RADseq data and ecological niche modeling, Nygaard et al. (2021) found that Carex jemtlandica and C. lepidocarpa in Carex sect. Ceratocystis represent separately evolving entities that remain distinct in sympatry despite ongoing hybridization and introgression. As such, we are moving toward solving the long-standing questions about taxon limits within this species complex, a problem evaluated multiple times using Sanger sequencing (Jiménez-Mejías et al., 2012, 2017) and allozyme variation (Hedrén & Prentice, 1996; Hedrén, 2002, 2004; Blackstock, 2007) without clear resolution. Population genomics using GBS, RADseq, or targeted sequencing methods offer hope to tackle many more species complexes in future, as has already been shown in previous research efforts on sedges (Escudero et al., 2014; Maguilla et al., 2017; Léveillé-Bourret et al., 2020; Luceño et al., 2021).
Recently, deeply sampled phylogenetic data sets of several species-rich Cyperaceae lineages have been the subject of both biogeographical and diversification analyses (e.g., Spalink et al., 2016a, 2016b; Martín-Bravo et al., 2019; Larridon et al., 2021c, 2021d). Here, two new studies help further understand biogeographical and diversification patterns in Cyperaceae. Benítez-Benítez et al. (2021) uncover the geographical vs. ecological diversification patterns in Carex sect. Phacocystis by integrating a phylogeny of 82% of extant species of this large group with GBIF occurrence data. Elliott et al. (2021) show that the genus Schoenus (Cyperaceae) is now monophyletic after recent taxonomic work in the last few years (Elliott & Muasya, 2017; Barrett et al., 2019; Barrett et al., 2020; Barrett et al., 2021b). Their study also indicates that Schoenus originated in the Western Australian biodiversity hotspot, from where it diversified into a lineage of >150 extant species dispersed around the globe.
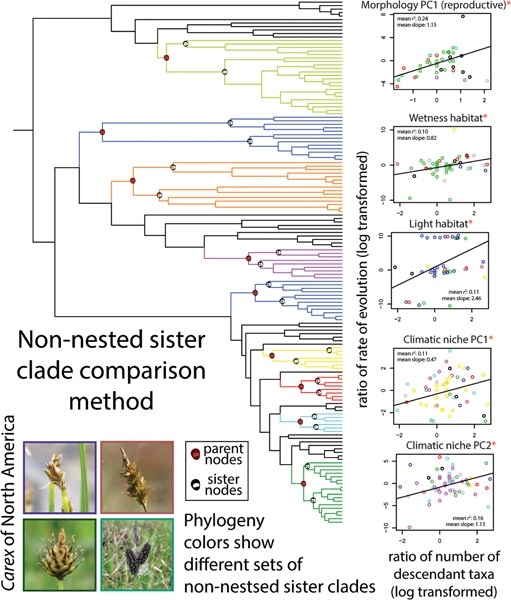
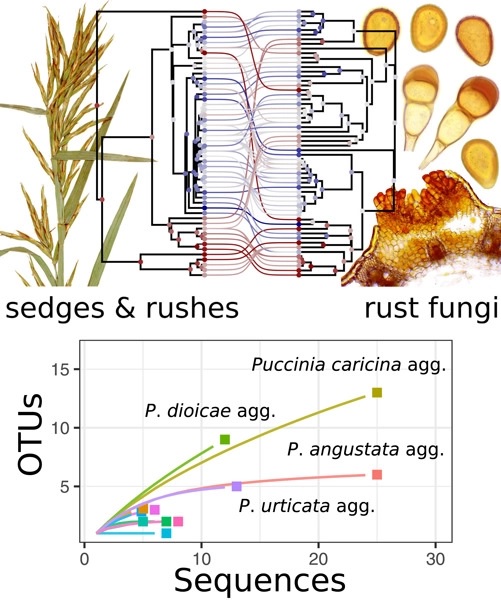
Large data sets also provide opportunities to address new questions. Here, Pender et al. (2021) show how trait evolution rates shape continental patterns of species richness in Carex, North America′s most diverse angiosperm genus, while Léveillé-Bourret et al. (2021) explore coevolution between parasitic rust fungi (Basidiomycota) and sedges and rushes. The data-rich era also allows for the Cyperaceae family to be used as a model family to investigate wider biological questions such as assessing the sensitivity of divergence time estimates to locus sampling, calibration points, and model priors (Villaverde et al., 2021), and assessing the effects of rapid chromosome number evolution on lineage diversification (Márquez-Corro et al., 2021).
We hope that this set of papers will stimulate future integrative research in sedges and promote the use of this extraordinary family as a model for exploring the diversity and evolution of life on Earth.